Il Pitone moluro bivittato nel Rettilario dello Zoo di Roma.
Accoppiamento e Deposizione
Nel Rettilario dello Zoo di Roma, sono state osservate fasi di corteggiamento, accoppiamento, deposizione e schiusa delle uova, in un periodo compreso fra il 1985 (anno del primo accoppiamento) ed il 1995, momento dell’ultima deposizione ottenuta nel corso di questa ricerca.
Si sono osservati i corteggiamenti fra i mesi di Novembre e Febbraio, perfettamente in sintonia con i dati forniti da Ross (1978). Le date di schiusa ed il numero dei nati ottenuti, hanno rivelato che la coppia osservata ha prodotto uova feconde dopo accoppiamenti avvenuti in Febbraio-Aprile, periodo riconosciuto come quello in cui invece avvengono la maggior parte delle schiuse in allevamenti dell’Emisfero Australe (Ross, 1978). Ritengo sia sempre stata la femmina ad andare in calore per prima, avendone osservato i caratteristici display descritti in letteratura (ancora non investigati approfonditamente). Il maschio sembrava inizialmente indifferente alle sollecitazioni fisiche ed odorose della partner, quali l’ondeggiamento e l’eversione della cloaca dinanzi a lui(Ross, 1978; Mehrtens, 1987). Il prolungarsi degli intervalli fra un pasto ed il successivo, hanno sempre accompagnato queste fasi pre-copula.
Il maschio di Pitone moluro impegna molto tempo nel rendere incline la femmina all’accoppiamento, passando e ripassando tutta la lunghezza del proprio corpo su quello della femmina e stimolando con pressioni e sfregamenti degli “speroni” sia il dorso che l’area della cloaca della partner. I corpi dei due pitoni non si avvolgono reciprocamente nella loro interezza, come si osserva in altre specie di serpenti (ad es. nei Generi Crotalus, Lampropeltis, Vipera, Coluber, Tamnophis, ecc.), ma il maschio, che assume una posizione sovrastante, avvolge la femmina con sole 2 o 3 spire in prossimita’ della cloaca, opponendovi la sua ed utilizzando gli speroni come ancoraggio supplementare per mantenere il contatto fra le due parti. In questa posizione, i Pitoni moluri possono restare per ore, talvolta muovendo entrambi la testa con piccoli scatti laterali. Questo e’ un movimento che il maschio effettua anche durante il corteggiamento, mentre si strofina sul corpo della femmina o mentre segue la scia odorosa della partner a pochi centimetri dalla sua cloaca ondeggiante. La coppia osservata allo Zoo di Roma e’ rimasta unita per oltre 5 ore in un singolo accoppiamento (pers. obs., 1994). I maschi di Pitone moluro non inducono all’accoppiamento una femmina afferrandola con la bocca in prossimita’ della nuca, come usano invece i maschi di altre specie, appartenenti a differenti generi (Coluber, Naja, Lampropeltis, etc.). Per tutto il periodo degli accoppiamenti, dal corteggiamento alla copula nei maschi e dalla gestazione (un mese circa prima della ovideposizione) alla schiusa delle uova nelle femmine, questi Ofidi non si alimentano. La ripresa dei ritmi di vita abituali, avviene lentamente e talvolta e’ necessario anche piu’ di un mese per tornare alla normalita’.
La femmina osservata nel Rettilario dello Zoo di Roma ha sempre deposto fra Maggio e Luglio (tab. ). Con l’approssimarsi del momento della deposizione, si e’ notato un cospicuo ingrossamento del terzo inferiore del corpo della femmina. Dopo essersi nutrita ancora per qualche tempo al termine dell’accoppiamento, ha iniziato ad ignorare cio’ che le veniva offerto in pasto (nel periodo degli accoppiamenti, solo il maschio e’ stato osservato non prestare alcuna attenzione al cibo). Nei successivi giorni, per un periodo di 10-15 giorni, e’ stata osservata sempre piu’ a lungo in prossimita’ del sito di deposizione, ove con movimenti concentrici delle spire spostando la sabbia del substrato, ha ricavato un avvallamento dalla forma circolare profondo non piu’ di 20 cm. e del diametro di 80-90cm. L’approssimazione di queste misure si giustifica col fatto che le modifiche a questa depressione prodotta sul terreno sono continue, anche dopo la deposizione. Si e’ inoltre notato che non sempre questa “tazza” viene totalmente utilizzata nella sua estensione, rappresentando anche un problema per la futura madre, quando il margine rialzato diviene un ostacolo se la femmina vuole allontanare dalle sue spire eventuali uova scartate per vari motivi dalla massa in cova (immature, rotte, attaccate da muffe ecc.ecc.).
Si e’ osservato la femmina rovesciare il corpo sul dorso; questo atteggiamento e’ sempre stato considerato dall’autore come “defaticante”, avendo notato l’aumento della frequenza del suo verificarsi al sopraggiungere del momento della deposizione e la sua totale assenza in qualsiasi altro periodo dell’anno.
 |
Femmina di P.m bivittatus in cova – Zoo di Roma 1987 |
Nonostante molti allevatori separino i sessi accostandoli nei soli mesi riproduttivi sino a copula avvenuta (in genere da Ottobre a Febbraio), nel Rettilario dello Zoo di Roma la coppia ospitata e’ sempre stata nel medesimo terrario per tutto l’anno.
In un lasso di tempo di 6-7 ore la femmina osservata, di 5m. e 55 kg. di peso, ha deposto nel Giugno 1992 54 uova, che ha immediatamente iniziato a covare, incrementando la sua temperatura corporea con le caratteristiche contrazioni delle spire del corpo. La temperatura e l’umidita’ del terrario sono state controllate quotidianamente dal momento della deposizione nei giorni successivi, curando di non far scendere i valori climatici al di sotto della norma: 29 C e 85% UR. Inoltre, per due volte, si e’ potuto misurare al 12° ed al 39° giorno di incubazione, la temperatura tra le spire della femmina, rilevando rispettivamente incrementi di 7 e 4 gradi C rispetto alla temperatura ambiente. Sembrerebbe esistere un rapporto diretto fra la frequenza delle contrazioni della femmina in cova e la temperatura ambiente. La letteratura propone intensita’ e ritmi di contrazioni caratteristici da individuo ad individuo (Paulraj e Thiruthalinathan, 1989). L’autore concorda sia con la tesi dell’esistenza di un rapporto diretto fra la frequenza delle contrazioni e la temperatura ambiente, sia con la tesi che propone una caratterizzazione individuale delle contrazioni. Una asserzione non esclude l’altra. Si e’ notato ripetutamente negli anni di riproduzioni, il calo della frequenza e della forza delle contrazioni avvicinandosi al momento della schiusa; cio’ non sarebbe casuale, ne’ caratteristico dell’individuo in osservazione.
Ci si chiede se gli embrioni abbiano bisogno di un apporto termico maggiore all’inizio del loro sviluppo. Le due femmine di Pitone moluro bivittato riprodottesi allo Zoo di Roma (l’altro esemplare osservato e’ una femmina nata nel 1986 dal soggetto di cui abbiamo trattato sin’ora), hanno sempre serrato con forza le spire in risposta alle sollecitazioni fisiche causate da chi scrive, sopratutto nei primi 20-30 giorni di cova, impedendo molte osservazioni e rilevamenti eseguiti invece qualche tempo dopo con relativa semplicita’. Le contrazioni immediatamente successive alla ovideposizione (un’ora dopo), erano intervallate da pause di 8-12 secondi l’una dall’altra; gia’ dal giorno successivo, questo spazio di tempo era salito a quasi 30 secondi.
 |
Femmina di P.m bivittatus Zoo di Roma 1987 |
Prendendo spunto da quanto riferito da Paulraj & Thiruthalinathan (1989), si e’ voluto verificare l’effettiva rotazione delle spire della femmina in cova attorno alle uova deposte e la sua entita’, marcando ogni spira del pitone e ponendo due altri punti di riferimento fra l’ofidio e l’osservatore. Utilizzando l’allineamento delle marcature effettuate sul pitone con una retta ideale passante per gli altri due punti (fissi), si e’ rilevata una irregolare rotazione del soggetto osservato, fatto non del tutto in accordo con quanto si evince dal lavoro di Paulraj & Thiruthalinathan. Inoltre, il senso di rotazione delle spire viene spesso invertito, ed anche questo sarebbe in contrasto con quanto sostenuto (Paulraj e Thiruthalinathan, 1989). Tale comportamento e’ interpretabile come il tentativo del pitone di distribuire omogeneamente calore e pressione esercitati sulle uova. Non si puo’ tuttavia escludere che vi sia anche il contemporaneo rimescolamento delle uova sottostanti, come semplice conseguenza fisica dei sussulti prodotti per generare il calore necessario alla cova (Paulraj e Thiruthalinathan, 1989). Quest’ultima considerazione trova riscontro in alcuni rilevamenti di controllo, eseguiti presso il Rettilario dello Zoo di Roma sulla femmina in cova nel Maggio 1995, lo stesso individuo tenuto in osservazione dal 1984, del quale abbiamo trattato sin’ora. Si sono riscontrati anche spostamenti laterali quasi quotidiani di alcuni centimetri (max. cm.23; min. cm.6) della femmina e della sottostante massa delle uova, rispetto all’asse perpendicolare al centro del nido (fig. n.1 ).
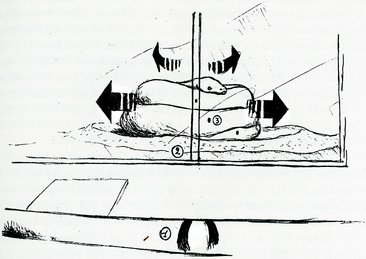 |
| Sistema di Allineamento a 3 punti per l’ osservazione dei movimenti effettuati dalla femmina di P.m. durante la cova. |